遗传学教学角度下对生物多样性的理解
我上遗传学
遗传与变异
爱你不容易
大家同显性
教学创新意
遗传学是研究生物遗传与变异规律的科学,是生命科学中最富有综合性的学科,也是最具时代气息的一门学科,正是由于这些特点,它赋予了遗传学教师充分的主观能动性和展示自我风采的机会。为了展现全国高校遗传学一线教师的教学风采,加强教师间的教学经验交流,让学生领略到不同知识点间的学习技巧,本公众号特邀全国一线遗传学教授,为大家奉上精彩的教学经验系列《我上遗传学》。
谈笑有鸿儒,往来无白丁。众人拾柴火焰高,相信同仁们的智慧能让我们的遗传学教学呈现异样的精彩!敬请关注。
前言
近代以来,随着科学技术的不断发展,人们认识自然以及改造自然的能力得到了大幅提升。但由于人类活动的加强,全球环境的剧烈改变,已经逐渐对生物多样性产生极大的影响,包括物种的行为、分布、丰富度、种群大小和种群关系等都已经发生了不同程度的影响,甚至引起某些物种的灭绝[1]。面对由生物多样性波动所造成的严重生态后果,人们不知不觉开始拉响了警钟。如何有效的保护生物多样性、维持各物种和生态系统的和谐发展已经成为国际社会普遍关注的重大问题。
遗传学是研究生物遗传与变异的科学,是生物学中一门十分重要的基础学科和指导育种工作的理论基础,同时也是探索生物多样性、生命起源和进化机制的重要理论基础。理解和保护生物多样性离不开遗传学理论及其技术的革新,同时对生物多样性的研究也补充和丰富了遗传学的内容。
遗传学的发展历经了经典遗传学、分子遗传学、基因组遗传学等多个阶段。本文以遗传学不同时期或不同研究水平的特点为视角,从遗传学教学的角度出发理解和认识生物多样性,企图丰富遗传学教学内涵,为遗传学教学提供参考。
生物多样性的概念
生物多样性是一个描述自然界生物多样性程度而内容广泛的概念。对于生物多样性,不同学者给出的概念有所不同,如蒋志刚等在《保护生物学》中的概念为:“生物多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的总和,包括动物、植物、微生物和它们拥有的基因以及它们与其生存环境形成的复杂生态系统,是生命系统的基本特征”[2];Wilson等人认为生物多样性是在特定的时刻和地点,生物体所呈现的种类和数量的丰富性[3];在联合国所制定的《生物多样性公约》中,生物多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的总和,这包括物种内部、物种之间和生态系统的多样性[4]。
生物多样性可以分为三个层次:(1)遗传多样性,广义上的遗传多样性是指地球上各种生物所携带的遗传信息的总和,狭义上的遗传多样性是指种内的遗传多样性,即种内个体之间或一个孟德尔群体内不同个体的遗传变异总和。(2)物种多样性,指生物群落中物种(动物、植物、微生物等)的丰富性和异质性。(3)生态系统多样性,即指生物圈内生境、生物群落和生态过程的多样化以及生态系统内生境差异、生态变化的多样性。
总之,生物多样性是由上述三个层次相互交结而成的复杂的生物学复合体系,其中,遗传多样性是物种多样性和生态系统多样性的基础,或者说遗传多样性是生物多样性的内在形式。物种多样性或生理表型多样性如繁殖方式的多样性、免疫系统的多样性等彰显了遗传的多样性,同样,生态系统多样性离不开物种的多样性,更离不开不同物种所具有的遗传多样性[5]。

图一:生物多样性()
经典遗传学水平上对生物多样性的理解
经典遗传学发展阶段主要指1953年以前的工作,在这一时期,建立了遗传学的三大基本定律,并确证了遗传的染色体学说和遗传的物质基础等。
自孟德尔的豌豆杂交实验开始,遗传学家们在自然繁殖规律中发现了生物多样性的存在,并开始交上自己的答卷。在经典遗传学中,运用“遗传因子”这一理论实体,借助孟德尔模型,抓住了待解释遗传现象中的主要因素即基因,从而大大简化了解释过程。人们开始借助遗传学这个有力的工具来认识自然,对生物多样性的认识也在此过程中不断得到发展。
经典遗传学中的三大定律为生物多样性的解释提供了理论基础:分离保证了遗传的稳定,自由组合和连锁交换产生了更多的基因组合。在减数分裂的配子形成过程中,成对的基因在杂种细胞中能够彼此互不干扰,独立分离,通过基因重组在子代继续表现各自的作用。分离定律从理论上说明了生物界由于杂交和分离所出现变异的普遍性。自由组合和连锁与交换定律解释了自然界生物发生变异的基本来源,二者的差别在于自由组合的基因是由非同源染色体所传递,而连锁交换的基因则是由同一对同源染色体所传递。,理论上,任何基因间或基因内都可以发生重组,再加上减数分裂中连锁与交换的必然性,其结果产生的生物多样性是难以想象的。从以上可以看出,经典遗传学三大定律已经充分说明了物种之间是如何孕育出如此丰富的生物多样性的了。
不同的物种享有不同的基因组成,构成不同的基因型,形成不同的表型性状,同一物种也会因染色体上不同的基因行为,导致性状差异,产生生物多样性。遗传学中的“一因多效”、“多因一效”、“表型模写”、“表现度”等同样可导致生物多样性的产生。等位基因间及非等位基因间的相互作用,除使孟德尔比例发生改变外,由于新类型的产生也导致生物多样性类型的产生,如基因互作产生新的生物类型等,这部分的内容在经典遗传学中称为上位效应(epistasis)[6]。基因突变是种内产生生物变异的重要基础来源,基因突变的结果导致等位基因(包括复等位基因)的产生,通常可引起一定的表型变化。而染色体畸变(包括染色体结构和数目变化)可对生物体的遗传信息造成更大的改变,产生新的表型效应甚至新物种的形成,极大地增加了遗传多样性。总而言之,基因的各种遗传行为是生物多样性的一个最主要的驱动力[7]。
利用经典遗传学原理发展了许多育种技术,如杂交育种、杂种优势创造、单倍体和多倍体育种、诱变育种等,除了在生产实践中发挥了巨大的作用外,如杂交育种产生的骡子同时具有驴的负重能力、抵抗能力和马的灵活性和奔跑能力,与此同时也带来了更丰富的生物多样性,如运用诱变育种技术,培育出了各种目标的农作物、家畜家禽以及园艺作物等。

图二:孟德尔遗传定律()
分子遗传学水平上对生物多样性的理解
自1953年沃森和克里克提出DNA分子双螺旋结构模型开始,遗传学进入分子遗传学阶段,从此使人们对生物多样性的认识更进一步。
DNA分子中不同的碱基对排列顺序,构成了不同的DNA组成。假设一个DNA分子中有n对碱基,它的排列方式便有4n种,实际上一个DNA分子含有成千上万对碱基,其排列方式几乎是无限的。正由于DNA分子的这种多样性,加上每种生物都有决定自己生物种质性状的特异性DNA序列,也就决定了生物的多样性。
分子遗传学除了让人们发现基因的分子世界蕴含着无穷的生物多样性基础外,也让人们看到了一种微观的、动态的生物多样性形成过程,从一段基因到其功能的实施,再到其最终表型的体现,中间涉及了大量基因与基因之间、蛋白与蛋白之间的相互作用。在漫长的物种演化过程中,为了自身更好的生存和繁衍,基因也在不断的发生新的演化以适应生物对环境的变化。例如,有研究认为藏族人选择性的EPAS1基因单体型来自于丹尼索瓦人或与之相似人群的古老基因,这一基因变异体的保留使藏族人能更好的适应高海拔地区的生活,从而避免大多数从别的地区迁移至高海拔地区人们常见的一些副作用如血液变得粘稠、导致心血管问题等[8]。
分子遗传学的灵魂是中心法则,遗传信息从DNA传递给RNA再传递给蛋白质。转录和翻译过程决定了多肽链中氨基酸的排列顺序,由多种氨基酸构成的蛋白质在生物体内执行不同的功能,引起一系列错综复杂的代谢变化,大部分分子水平的变异都会通过影响上述过程,展现出个体之间细胞、器官、生理代谢以及形态学水平的差异。分子水平的基因变异可以是由于DNA分子中发生碱基对的增添、缺失或替换而引起的基因突变,也可以是在中心法则实现过程中,由于生物体受内外环境因素影响,发生选择性剪接、RNA编辑、表观遗传修饰等差别而造成生物多样性[9]。例如,人体的肤色是一种受基因与环境共同影响的复杂性状,多数相关基因控制黑色素的合成、转运及降解途径来控制肤色,当黑色素生成酶的相关基因发生突变或某个核苷酸发生多态性改变时,有关黑色素的相关过程均会受到影响,从而导致人与人之间肤色的差异[10]。有研究发现非洲人群的MC1R(黑皮质素1受体基因)序列变异较少,而欧洲人群却变异较多[11]。
通过体外DNA重组技术和转基因技术,赋予生物以新的遗传特性,创造出更符合人们需要的生物类型和生物产品,这一基因工程技术是分子遗传学应用最突出的成就,已显示出广阔的应用前景。转基因育种技术的发展,不仅打破了基因的物种界线,产生了新的生物品种,也扩宽了生物的多样性。当然,这也引发了更多人类对自然干预的担忧。

图三:中心法则()
基因组遗传学水平上对生物多样性的理解
1990年人类基因组计划的正式启动,使人们迎来了一个崭新的生物学时代--基因组时代,同时也使遗传学的研究进入一个新时代--基因组遗传学时代。
任何一个物种都具有其独特的基因库和遗传组织形式,基因组的多样性是不同物种间彰显遗传多样性的直接原因。很早人们就从等位酶的存在,意识到物种间的分子多样性具有较大差异,后来在基因组层面上得到证实。例如,果蝇有4对染色体 共有1万至1.5万个基因,基因组中有大约3%的位点是可变的[12],人类基因组由23对染色体组成,约有30亿个碱基对,其中只有0.1%显示等位基因变异[13]。基因的多态性在不同基因位点和染色体区域也存在较大差异,这在大量不同的植物、动物、真菌中均得到证明。从某种程度上说,遗传多样性可以被认为是在基因层面上遗传变异(等位基因)出现与消失所达到的一个平衡,新一代物种的产生会带来新的等位基因。虽然DNA中的大多数位置可以随机变化多次,但某些具有重要功能的位点在很长时间(如1亿年内)都没有发生变化,从这点上可以在一定程度上预测其进化的方向和潜力[14]。

图四:每个基因组变异位点的标准化数量[13]
(Auton et al.,2015)
在基因组遗传学时代,传统地用简单的孟德尔遗传规律以及“一基因一酶一功能”的假设来认知基因的功能是不可行的,科学家们开始从宏观的角度出发去研究基因,蛋白及其分子间的相互作用,利用全基因数据可以获得更多的基因组信息,从而进一步阐释遗传多样性目前的数量与结构,以及遗传多样性的起源。目前在猪、牛、羊等畜禽中不断开展广泛系统的全基因组重测序研究,且已取得了许多不错的进展[15],Medugorac等利用基因芯片分别对蒙古牦牛和家养牦牛进行重基因组测序,发现牦牛基因组中携带了一部分的黄牛基因组,且牦牛的无角性状来自蒙古牛[16]。通过广泛的基因组比较,可以研究不同物种如何适应特定环境,为物种所呈现遗传多样性的结构做出一定解释。例如,基因型范围较窄且表型更为一致的种群可能更倾向于在局部环境中变得不那么有利,在极端情况下,这样的种群更有可能灭绝[17],大熊猫之所以成为国宝,就在于它的低遗传多样性水平所导致的对自然环境压力的适应能力较差所致。
基因组遗传学时代也同样带来了新的、高效的育种方式。基因组选配可以在动物选育中控制ΔF(近交水平)增长时获得最大化ΔG(遗传进展)或找到两者间的平衡点,同时提高育种的质量和效率[18]。越来越多分子标记的发现,已被广泛应用于生物多样性的研究如个体间亲缘关系鉴定、种群遗传分析、遗传作图等以及分子标记育种。基因组编辑技术的应用能更进一步克服转基因中的一些弊端,如整合位点更准确、生命伦理问题更少等[19]。
总之,随着组学时代相关学科技术的发展与应用,现代生命科学的理论框架不断丰盈,这使我们可以从整体的角度、不同的层面上更加了解生命体之间相互作用的机制,更好的把握生物多样性的起源、发展与进化等。
表观遗传学水平上对生物多样性的理解
现代遗传学研究表明,基因表达模式在细胞世代间的可遗传性并不完全取决于基因的序列信息,这种不涉及DNA序列改变的基因表达和调控的可遗传变化称为表观遗传(epigenetic)。环境因素对表观遗传有着重要的影响,如所摄取的营养、所暴露的环境等都会影响基因的表达和表达模式的改变,从而产生表型的改变。表观遗传变异同样可以产生包括形态特征、生理特征、行为特征、甚至性别变异等表型变异,这对生物多样性同样产生巨大的作用。当然,若在生物发育过程中,环境压力源诱导的表观遗传变异产生不适应表型,同样可能对种群的演化产生影响。
表观遗传学使人们认识到,同基因组的序列一样,表观遗传修饰也包含有遗传信息,如DNA甲基化、染色质重塑、组蛋白修饰、基因组印记、非编码RNA等,尤其是DNA甲基化,代表了生物多样性的一个分子组成部分,它直接将基因组与环境联系起来。DNA甲基化对环境非常敏感,它参与了生物体对环境变化的可塑性和适应性反应,从影响个体表型变异到生态系统水平的所有生物的生态和进化过程。例如,一些鱼类和爬行动物的环境性别决定(environmental sex determination, ESD)主要依赖于cyp19a1基因的表达(该基因编码参与卵巢分化的芳香化酶),在不同环境的驱动下会引起其启动子甲基化状态的改变从而影响该基因的表达[20]。一些学者认为,鉴于当前全球变暖,这种表观遗传介导的ESD可能通过改变自然种群的性别比进而影响种群结构,造成生物多样性的波动[21]。
不同物种的表观遗传机制存在较大差别,譬如说,不同物种之间DNA甲基化的模式和水平存在差异。例如,在脊椎动物中,生物体通常表现出高水平的甲基化,以连续的方式分布在整个基因组中,通常与活性基因的启动子和调控序列相对应[22]。这些特定基因组区域的甲基化通常会抑制相关基因的转录,从而最终影响细胞和生物体的表型;脊椎动物早期发育过程中体细胞的特殊化需要对DNA甲基化模式进行广泛的清除和再编程,而植物则完整地将上一代的甲基化传递给下一代,在一些鱼类(如斑马鱼)中,DNA甲基化的清除只发生在雌性性腺发育期间,而在雄性性腺中则保持[23]。这说明雄性的DNA甲基化模式可能受到环境因素的影响,至少部分会传递给下一代。在鸟类、两栖动物和爬行动物中,DNA甲基化通常也以连续的方式分布在基因组中,很少存在DNA甲基化再编程现象。无脊椎动物基因组的甲基化水平明显低于脊椎动物,甚至在一些无脊椎动物物种中,未检测到DNA甲基化(例如秀丽隐杆线虫)或只有极低水平的DNA甲基化(小于基因组的1%;例如果蝇),这清楚地表明DNA甲基化并不是这些物种发育的关键因素[22]。不同于哺乳动物基因组只有CG甲基化,植物基因组甲基化有CG,CHG,CHH(H代表任何非G的碱基)甲基化[24]。而维持这三种不同的DNA甲基化的分子机制非常复杂,毕竟,植物基因组本身比较庞大,且很多物种是多倍体的,加上有不断跳跃的转座子,还需要应对复杂的环境变化,通过甲基化让自身基因组变得安分,似乎也是比较明智的选择。
结语
从某种角度上来说,生物多样性是动态的、变化的遗传事件的反映。遗传学研究虽然经历了一个多世纪的时间,我们也比以往任何时候都更了解生命的多样性及其巨大的进化史,并且不断开拓生物多样性新的研究内容。然而,我们仍然只看到了生物复杂性的冰山一角。正如Sandell和Otto在思考《Genetics》杂志在其第二个世纪将要刊载的内容时所认为的:“我们对遗传学的看法将逐渐从试图理解生物学的一般模式转向设法解决生物学的多样性,这样做可以更好地揭示生物学复杂性的深度”[25]。
过去的遗传学集中于解释基因的主要影响以及共同的遗传模式,而现在的遗传学试图去解释基因的功能差异、产生生物多样性的原因以及生物进化的路线差异等,揭示越来越多的物种、基因在不同环境中不同寻常的作用。这也就意味着站在遗传学的视角,展望生物多样性,可以让我们更好的感受到地球这个丰富多彩大生物圈的奥妙之所在,甚至预期人类及其生物圈的未来世界。
生物多样性是生物历经遗传、变异、进化漫漫征途的必然结果。每一生物个体都是基因的载体,生物的多样性代表着41亿年来生命演化的结果[26],对于人类来说是一笔巨大的财富,为我们提供氧气、食物、药物以及生命活动所必须的物质和能量。任何一个物种的灭绝,,对人类来说将是永远的损耗,所以更好的理解和保护生物多样性,防止物种灭绝是我们人类的责任和义务。在遗传学教学中深刻领会生物多样性的丰富多彩及其机制,对于遗传学的教学内涵的挖掘及生物多样性的保护和地球物种的生生不息具有重大的战略意义。
相关阅读
-

在日本人眼中世界上科技实力最强的10座城市看都有哪些...
-

2023济南历城区哪里有装配式工程师培训学校(有关装配式工程师的政策)
2023济南历城区哪里有装配式工程师培训学校,人每天要吃东西,补充能量才能抵抗消耗。大脑需要不断学习才能补充新知识,今天学习内容看一下济南装配式工程师培训...
-
![[明瀚妇科]宫颈息肉是否需要治疗?最可怕的后果是这个!](https://www.lwdmc.com/uploadfile/thumb/20231220/1703044614405_2.jpg)
如不及时治疗,息肉会逐渐长大,可能会影响怀孕或造成性交出血。抛开这些因素不谈,更为严重的是,宫颈息肉具有一定的癌变倾向。因此...
-

该研究通过对实验室大鲵幼体胃肠道微生物群落的分析,揭示了环境温度可能对微生物群落有间接影响。...
-
南昌装配式工程师学习去哪家培训学校,想知道南昌装配式工程师培训。就了解下基本知识,什么是装配式工程师,装配式工程师线上考试要求,装配式工程师发展前景,装配式工程师就业方向...
-
2023济南历城区哪里有装配式工程师培训学校(有关装配式工程师的政策)
2023济南历城区哪里有装配式工程师培训学校,人每天要吃东西,补充能量才能抵抗消耗。大脑需要不断学习才能补充新知识,今天学习内容看一下济南装配式工程师培训...
-
摘要:【目的】 评价聚乳酸(polylactic acid,PLA)材料在不同土壤环境中自然降解的效果,通过对3种不同土壤菌群结构的分析,找到能够对聚乳酸材料有降解作用的优势......
-

全球科研城市50强武汉排19位 国内第4位 紧跟北京上海南京
武汉晨报讯(记者李佳)11月1日,最新出版的英国《自然》增刊“2018自然指数-科研城市”中,全球科研城市50强中有10座中国城市入选,武汉排19位。...
-
教改理论018年5月5一、年少不知天高地厚记得1989年9月,我从昆明师范学校毕业,被石林县教育局分派到板桥乡青山口完小教书。那时的我年轻气盛...
-
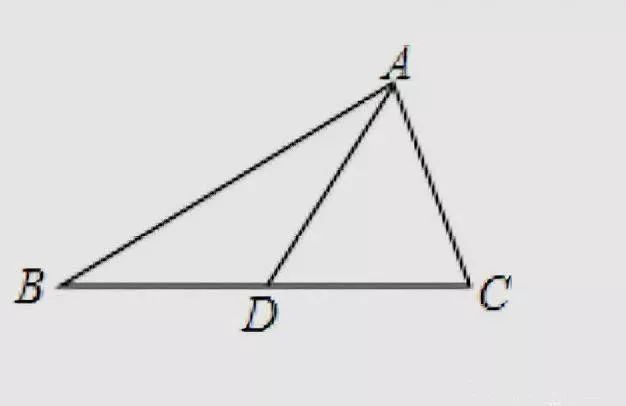
shuxueds如图所示,在△ABC中,AB=8,AC=6,AD是△ABC的中线,则△ABD与△ADC的周长之差为多少。...
发表评论
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任如发现本站有涉嫌抄袭侵权/违法违规的内容,请发送邮件举报,一经查实,本站将立刻删除。
